
Molecular Markers in Maternal Blood Exosomes Allow Early Detection of Fetal Alcohol Spectrum Disorders
 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
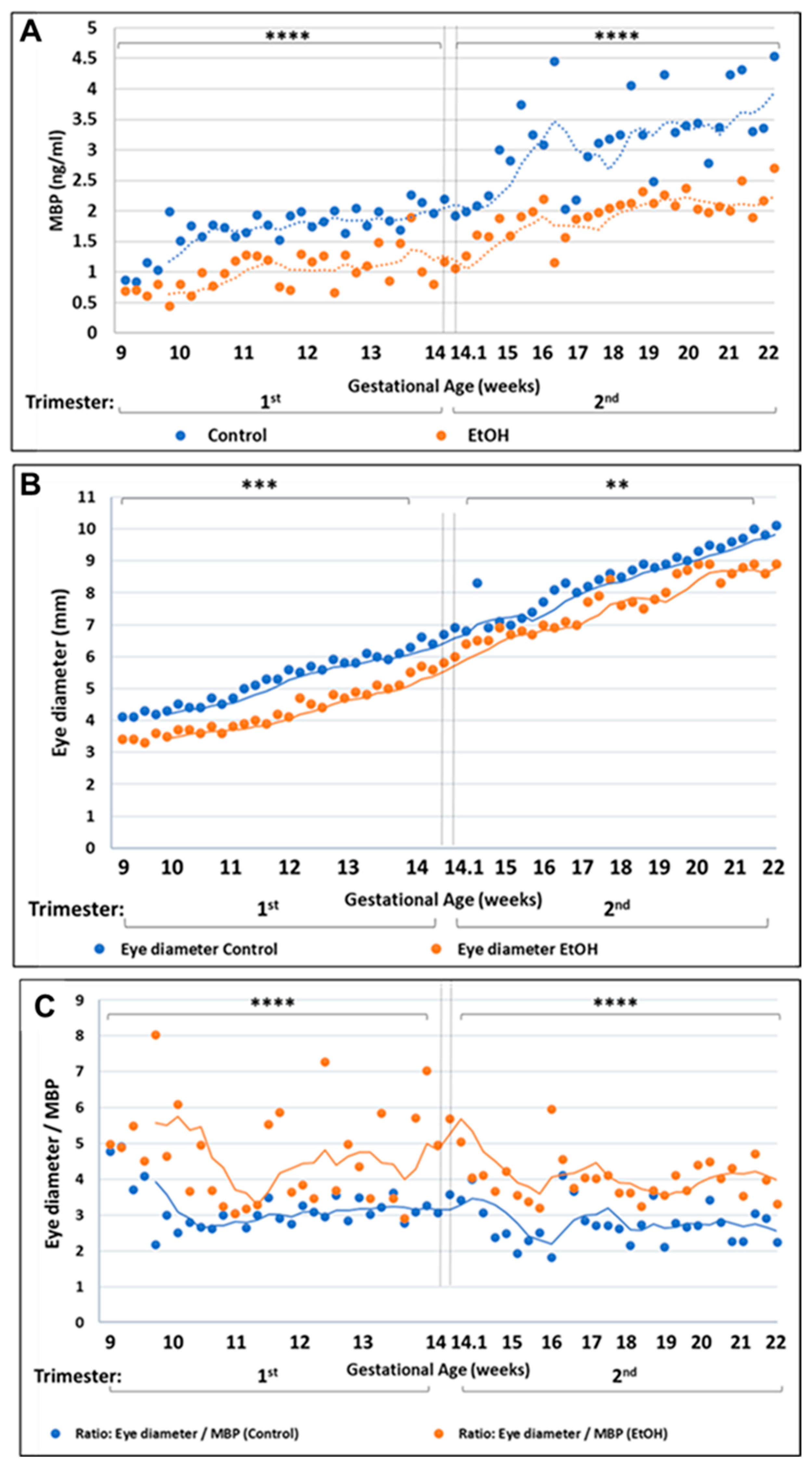
2.1. Prenatal EtOH Exposure Inhibits Eye Development in Human Fetuses
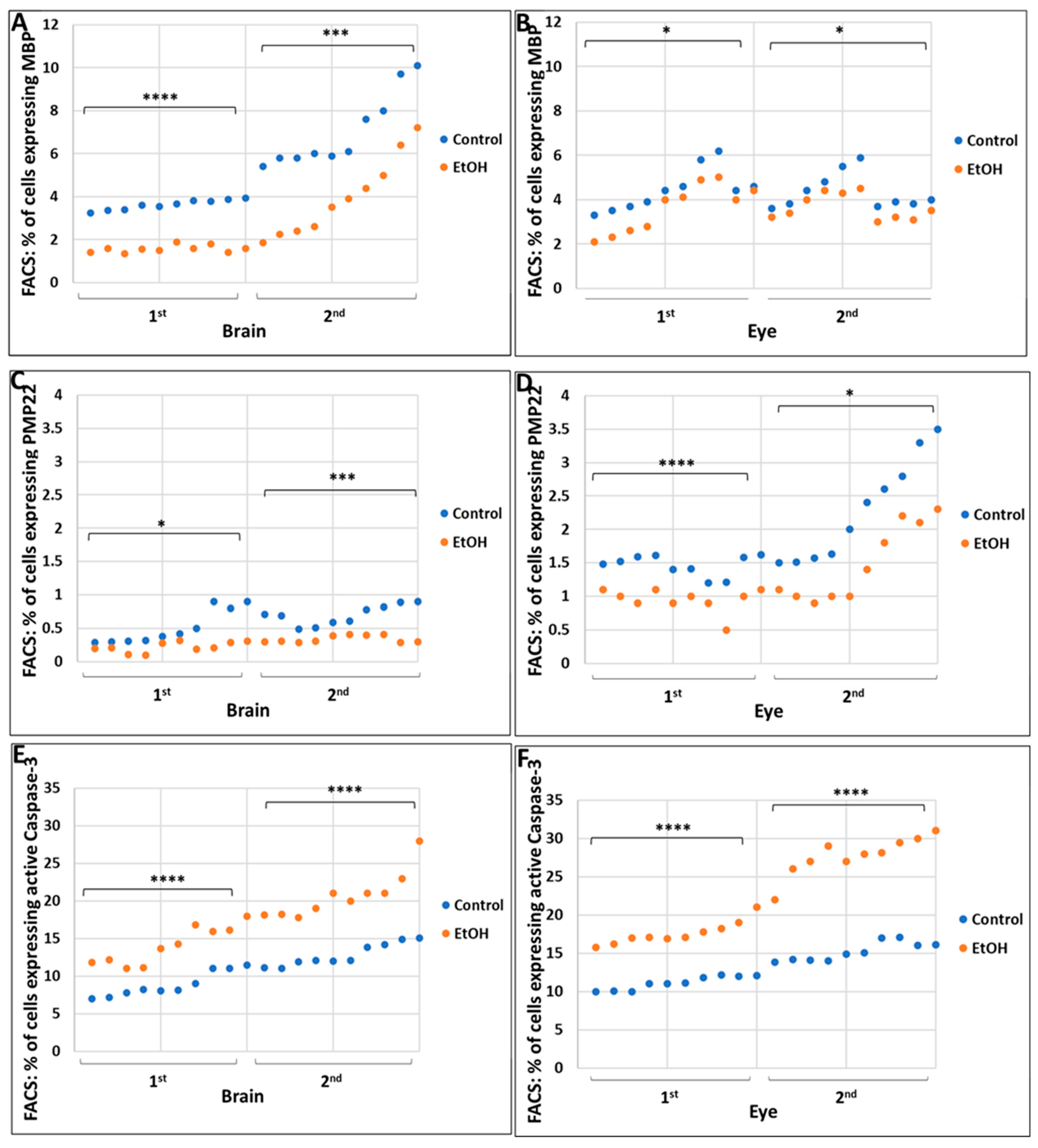
2.2. Prenatal Alcohol Exposure Inhibits Expression of the CNS and PNS Myelination Markers Myelin Basic Protein (MBP) and Peripheral Myelin Protein-22 (PMP22), Respectively, and Increases Expression of the Apoptosis Marker Activated Caspase-3, during Eye and Brain Development
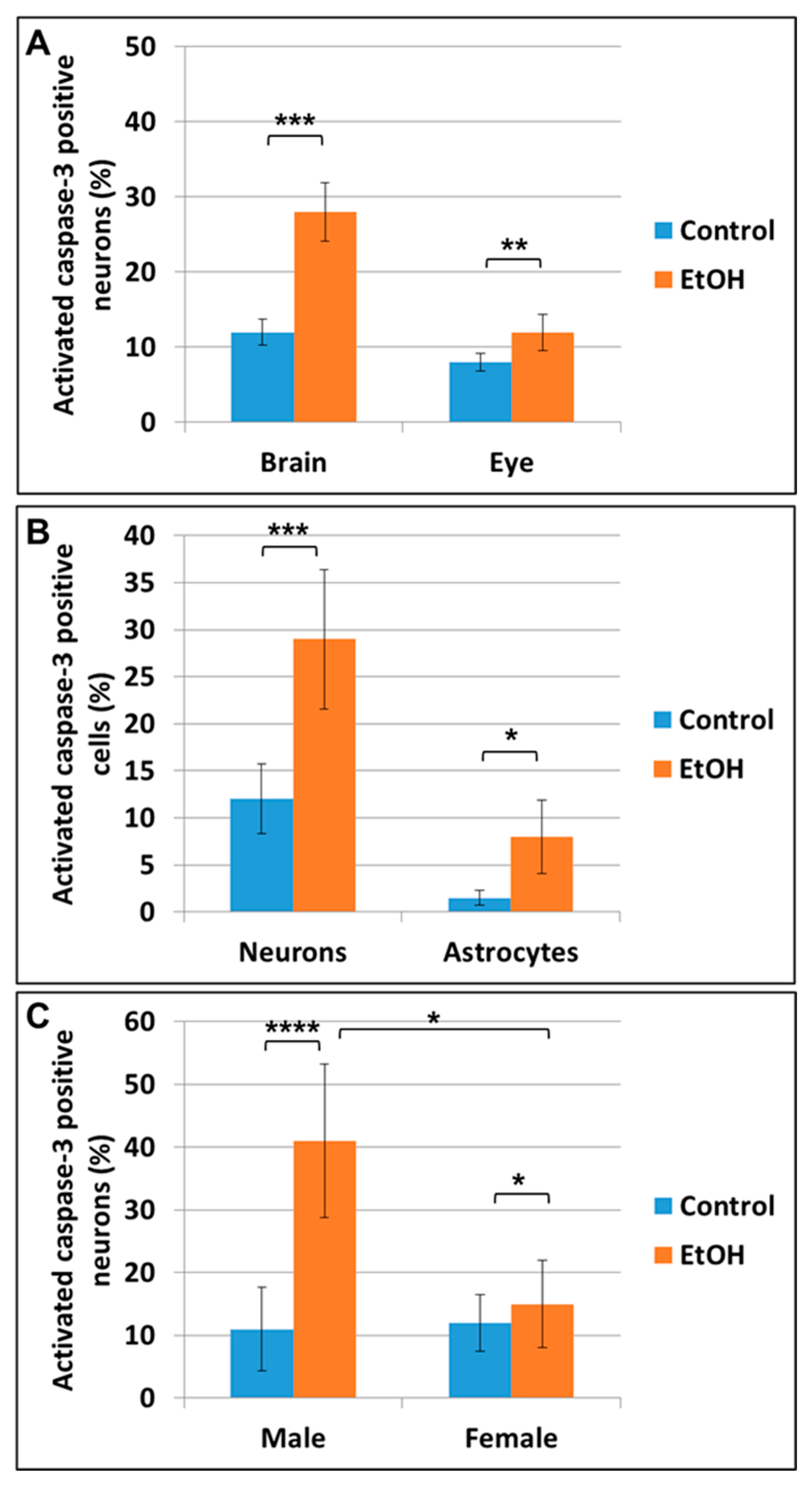
2.3. Prenatal EtOH Exposure Induces Neuronal and Astrocytic Injury in Fetal Brain and Eye
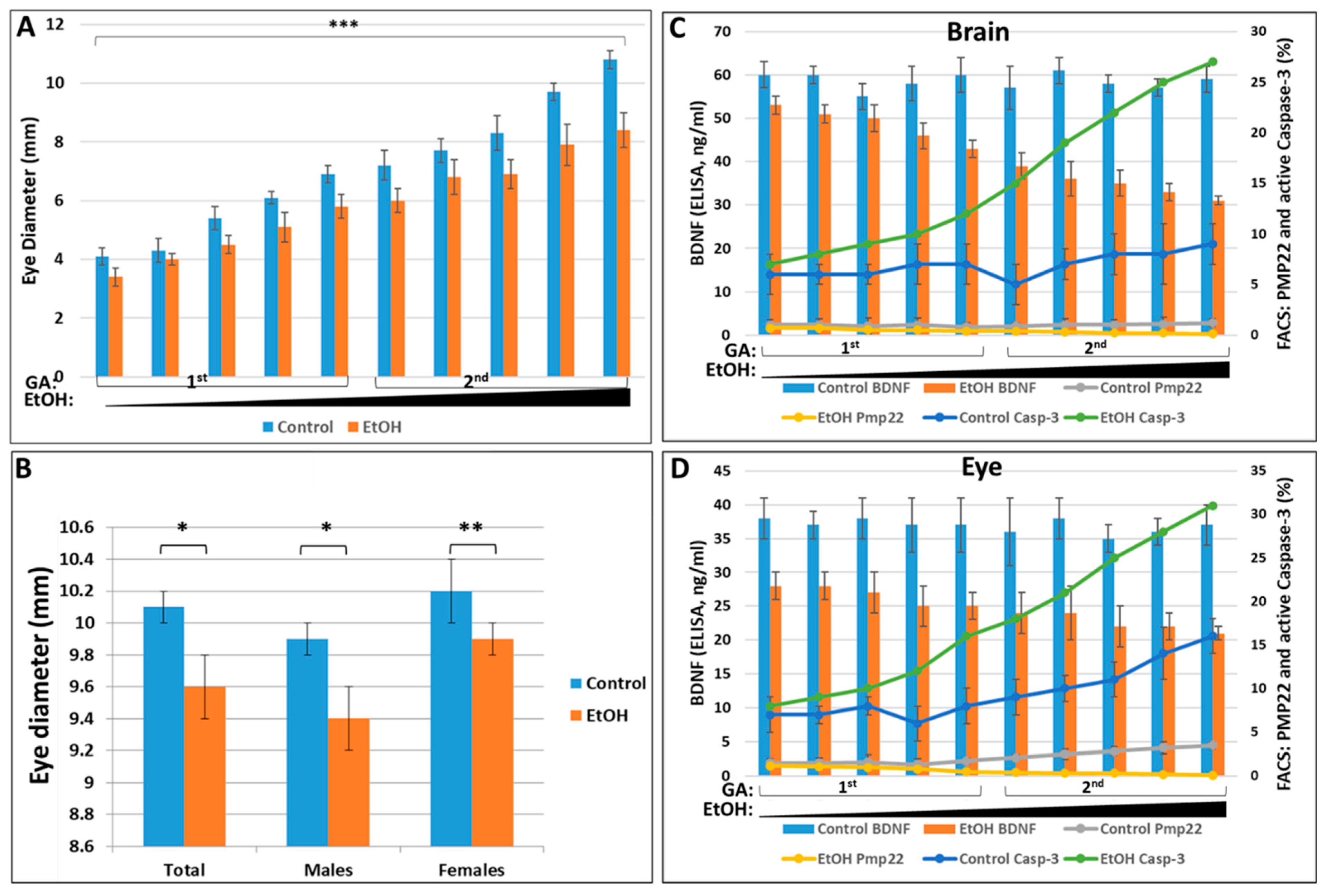
2.4. Prenatal EtOH Exposure Inhibits BDNF mRNA and Protein Expression in Fetal Eye, Brain and Synaptosomes
2.5. EtOH Dose, Gender, and Race All Strongly Affect Fetal Eye Development
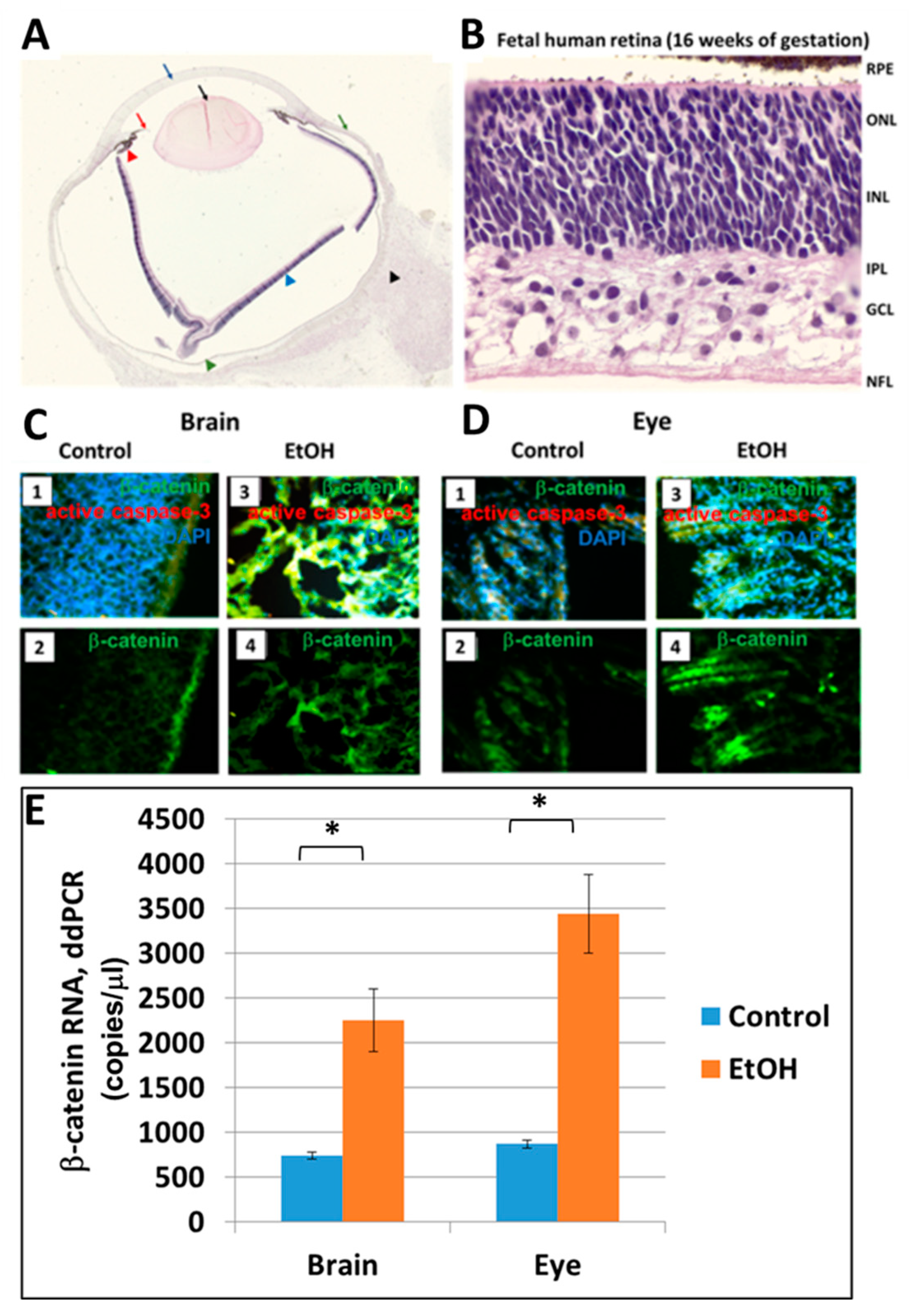
2.6. Prenatal EtOH Exposure Upregulates β-Catenin in Brain and Eye Tissues
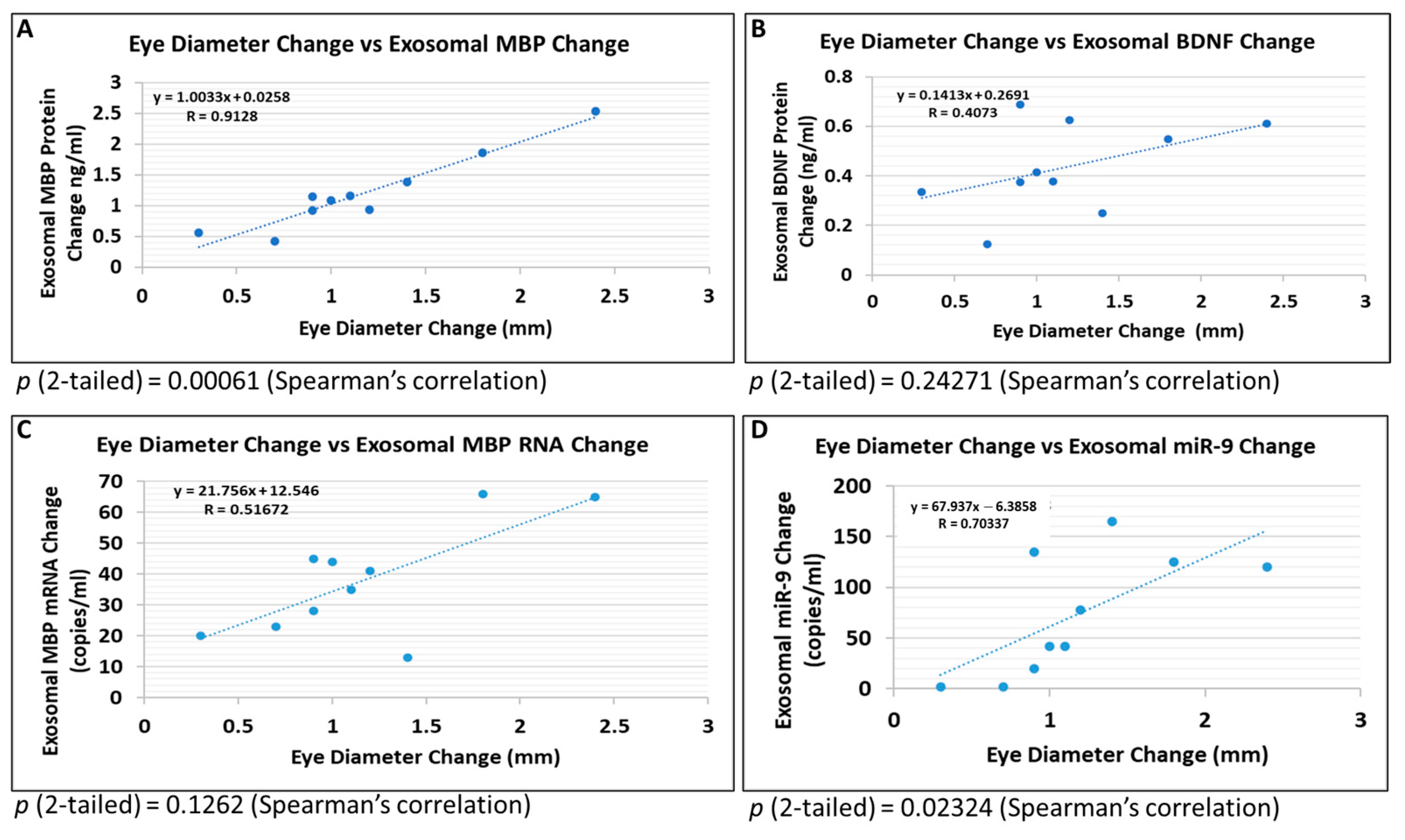
2.7. Brain-Derived Exosomes Provide Biomarkers to Diagnose Early FASD in Fetuses
3. Discussion
3.1. Reduced Eye Size as a Morphological Indicator of Alcohol-Associated Fetal Pathology
3.2. Potential Molecular Biomarkers
4. Methods
4.1. Clinical Samples
4.2. RNA Preparation and Real-Time Quantitative Polymerase Chain Reaction (qRT-PCR)
4.3. Sex Determination using Human Fetal Genomic DNA
4.4. Droplet Digital PCR (ddPCR)
4.5. Primers (IDT Inc.)
4.6. Flow Cytometry
4.7. ELISA Quantification of Brain, Eye, Synaptosomal and Exosomal Proteins
4.8. Immunohistochemistry
4.9. Antibodies
4.10. Isolation of Fetal Brain-Derived Exosomes (FB-Es) or Fetal OL-Derived Exosomes (OL-Es) from Maternal Plasma, and ELISA Quantification of Exosomal Proteins
4.11. Statistical Analysis
4.12. Ethics, Human Subjects
- a.
- Eligibility Criteria: The blood and tissue samples were obtained according to NIH Guidelines through a trained Study Coordinator. Samples were collected regardless of sex, ethnic background, and race.
- b.
- Treatment Plan: Each patient was asked to sign a separate consent form for research on blood and tissue samples. Blood obtained was processed for collection of serum and plasma. No invasive procedures were performed on the mother, other than those used in her routine medical care. Fetal tissues were processed for RNA or protein isolation.
- c.
- Risk and Benefits: There are very small risks of loss of privacy as with any research study where protected health information is viewed. The samples were depersonalized before they were sent to the lab for analysis. There were no additional risks of blood sampling as this was only performed in patients with clinically indicated venous access. There was little anticipated risk from obtaining approximately 2–3 cc of blood, but a well-trained Study Coordinator collected all samples.
- d.
- Informed Consent: Consent forms were maintained by the Study Coordinator and were not sent to the investigator with the samples. The de-identified log sheets and IRB protocol were sent by the Study Coordinator to Principal Investigator with each blood and tissue sample. This sheet contains an assigned accession number, the age, sex, ethnicity, and race of the patient. Except for an assigned accession number, no identification was kept on the blood and tissue samples.
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ornoy, A.; Ergaz, Z. Alcohol Abuse in Pregnant Women: Effects on the Fetus and Newborn, Mode of Action and Maternal Treatment. Int. J. Environ. Res. Public Health 2010, 7, 364–379. Available online: https://www.mdpi.com/1660-4601/7/2/364/htm (accessed on 21 April 2022). [CrossRef] [PubMed] [Green Version]
- Centers for Disease Control and Prevention (CDC). Alcohol use and binge drinking among women of childbearing age—United States, 2006–2010. MMWR Morb. Mortal. Wkly. Rep. 2012, 61, 534–538. Available online: https://pubmed.ncbi.nlm.nih.gov/22810267/ (accessed on 22 March 2022).
- Mutch, R.C.; Jones, H.M.; Bower, C.; Watkins, R.E. Fetal Alcohol Spectrum Disorders: Using Knowledge, Attitudes and Practice of Justice Professionals to Support their Educational Needs. J. Popul. Ther. Clin. Pharmacol. J. Ther. Popul. Pharmacol. Clin. 2016, 23, e77–e89. Available online: https://pubmed.ncbi.nlm.nih.gov/27132254/ (accessed on 22 March 2022).
- Hoyme, H.E.; Kalberg, W.O.; Elliott, A.J.; Blankenship, J.; Buckley, D.; Marais, A.S.; Manning, M.A.; Robinson, L.K.; Adam, M.P.; Abdul-Rahman, O.; et al. Updated Clinical Guidelines for Diagnosing Fetal Alcohol Spectrum Disorders. Pediatrics 2016, 138, e20154256. Available online: https://pediatrics.aappublications.org/content/138/2/e20154256 (accessed on 1 August 2016). [CrossRef] [Green Version]
- Sampson, P.D.; Streissguth, A.P.; Bookstein, F.L.; Little, R.E.; Clarren, S.K.; Dehaene, P.; Hanson, J.W.; Graham, J.M., Jr. Incidence of fetal alcohol syndrome and prevalence of alcohol-related neurodevelopmental disorder. Teratology 1997, 56, 317–326. [Google Scholar] [CrossRef]
- Andersen, A.-M.N.; Andersen, P.K.; Olsen, J.; Gronbaek, M.; Strandberg-Larsen, K. Moderate alcohol intake during pregnancy and risk of fetal death. Int. J. Epidemiol. 2012, 41, 405–413. [Google Scholar] [CrossRef] [Green Version]
- Popova, S.; Lange, S.; Probst, C.; Gmel, G.; Rehm, J. Estimation of national, regional, and global prevalence of alcohol use during pregnancy and fetal alcohol syndrome: A systematic review and meta-analysis. Lancet Glob. Health 2017, 5, e290–e299. Available online: https://www.thelancet.com/journals/langlo/article/PIIS2214-109X(17)30021-9/fulltext (accessed on 1 March 2017). [CrossRef] [Green Version]
- May, P.A.; Tabachnick, B.; Hasken, J.M.; Marais, A.S.; de Vries, M.M.; Barnard, R.; Joubert, B.; Cloete, M.; Botha, I.; Kalberg, W.O.; et al. Who is most affected by prenatal alcohol exposure: Boys or girls? Drug Alcohol Depend. 2017, 177, 258–267. Available online: https://pubmed.ncbi.nlm.nih.gov/28624747/ (accessed on 1 August 2017). [CrossRef]
- Abbott, C.W.; Kozanian, O.O.; Kanaan, J.; Wendel, K.M.; Huffman, K.J. The Impact of Prenatal Ethanol Exposure on Neuroanatomical and Behavioral Development in Mice. Alcohol. Clin. Exp. Res. 2016, 40, 122–133. Available online: https://pubmed.ncbi.nlm.nih.gov/26727530/ (accessed on 1 January 2016). [CrossRef] [Green Version]
- Donald, K.A.; Eastman, E.; Howells, F.M.; Adnams, C.; Riley, E.P.; Woods, R.P.; Narr, K.L.; Stein, D.J. Neuroimaging effects of prenatal alcohol exposure on the developing human brain: A magnetic resonance imaging review. Acta Neuropsychiatr. 2015, 27, 251–269. [Google Scholar] [CrossRef] [Green Version]
- Hanson, J.W.; Jones, K.L.; Smith, D.W. Fetal alcohol syndrome. Experience with 41 patients. JAMA 1976, 235, 1458–1460. Available online: https://pubmed.ncbi.nlm.nih.gov/946444/ (accessed on 22 March 2022). [CrossRef] [PubMed]
- González, E.R. New ophthalmic findings in fetal alcohol syndrome. JAMA J. Am. Med. Assoc. 1981, 245, 108. [Google Scholar] [CrossRef] [PubMed]
- Strömland, K.; Pinazo-Durán, M.D. Ophthalmic involvement in the fetal alcohol syndrome: Clinical and animal model studies. Alcohol Alcohol. 2002, 37, 2–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribeiro, I.M.; Vale, P.J.; Tenedorio, P.A.; Rodrigues, P.A.; Bilhoto, M.A.; Pereira, H.C. Ocular manifestations in fetal alcohol syndrome. Eur. J. Ophthalmol. 2007, 17, 104–109. Available online: https://pubmed.ncbi.nlm.nih.gov/17294389/ (accessed on 23 March 2022). [CrossRef] [PubMed]
- Sawada, K.; Sakata-Haga, H.; Komatsu, S.; Ohta, K.; Jeong, Y.-G.; Fukui, Y. A selective loss of small-diameter myelinated optic nerve axons in rats prenatally exposed to ethanol. Congenit. Anom. 2002, 42, 125–129. Available online: https://pubmed.ncbi.nlm.nih.gov/12196709/ (accessed on 23 March 2022). [CrossRef]
- West, J.R. Fetal alcohol-induced brain damage and the problem of determining temporal vulnerability: A review. Alcohol Drug Res. 1987, 7, 423–441. Available online: https://pubmed.ncbi.nlm.nih.gov/3304313/ (accessed on 23 March 2022).
- West, J.R. Recent findings on the mechanisms by which alcohol damages the developing nervous system. Alcohol Alcohol. (Oxf. Oxfs.) Suppl. 1994, 2, 395–399. Available online: https://pubmed.ncbi.nlm.nih.gov/8974361/ (accessed on 23 March 2022).
- Luo, J. Growth factor-mediated neural proliferation: Target of ethanol toxicity. Brain Res. Rev. 1998, 27, 157–167. [Google Scholar] [CrossRef]
- Ikonomidou, C.; Bittigau, P.; Ishimaru, M.J.; Wozniak, D.F.; Koch, C.; Genz, K.; Price, M.T.; Stefovska, V.; Horster, F.; Tenkova, T.; et al. Ethanol-induced apoptotic neurodegeneration and fetal alcohol syndrome. Science 2000, 287, 1056–1060. Available online: https://www.ncbi.nlm.nih.gov/pubmed/10669420 (accessed on 15 April 2022). [CrossRef]
- Goodlett, C.R.; Horn, K.H. Mechanisms of alcohol-induced damage to the developing nervous system. Alcohol Res. Health J. Natl. Inst. Alcohol Abus. Alcohol. 2001, 25, 175–184. Available online: https://pubmed.ncbi.nlm.nih.gov/11810955/ (accessed on 22 March 2022).
- Guerri, C. Glia and Fetal Alcohol Syndrome. NeuroToxicology 2001, 22, 593–599. [Google Scholar] [CrossRef]
- Guerri, C.; Bazinet, A.; Riley, E.P. Foetal Alcohol Spectrum Disorders and Alterations in Brain and Behaviour. Alcohol Alcohol. 2009, 44, 108–114. [Google Scholar] [CrossRef] [Green Version]
- Olney, J.W. New Insights and New Issues in Developmental Neurotoxicology. NeuroToxicology 2002, 23, 659–668. [Google Scholar] [CrossRef]
- Leigland, L.A.; Ford, M.M.; Lerch, J.P.; Kroenke, C.D. The influence of fetal ethanol exposure on subsequent development of the cerebral cortex as revealed by magnetic resonance imaging. Alcohol. Clin. Exp. Res. 2013, 37, 924–932. Available online: https://pubmed.ncbi.nlm.nih.gov/23442156/ (accessed on 1 June 2013). [CrossRef] [Green Version]
- Newville, J.; Valenzuela, C.F.; Li, L.; Jantzie, L.L.; Cunningham, L.A. Acute oligodendrocyte loss with persistent white matter injury in a third trimester equivalent mouse model of fetal alcohol spectrum disorder. Glia 2017, 65, 1317–1332. [Google Scholar] [CrossRef]
- Creeley, C.E.; Dikranian, K.T.; Johnson, S.A.; Farber, N.B.; Olney, J.W. Alcohol-induced apoptosis of oligodendrocytes in the fetal macaque brain. Acta Neuropathol. Commun. 2013, 1, 23. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Cuzon Carlson, V.C.; Studholme, C.; Newman, N.; Ford, M.M.; Grant, K.A.; Kroenke, C.D. In utero MRI identifies consequences of early-gestation alcohol drinking on fetal brain development in rhesus macaques. Proc. Natl. Acad. Sci. USA 2020, 117, 10035–10044. Available online: https://www.pnas.org/content/117/18/10035 (accessed on 25 May 2022). [CrossRef]
- Darbinian, N.; Darbinyan, A.; Merabova, N.; Bajwa, A.; Tatevosian, G.; Martirosyan, D.; Zhao, H.; Selzer, M.E.; Goetzl, L. Ethanol-mediated alterations in oligodendrocyte differentiation in the developing brain. Neurobiol. Dis. 2021, 148, 105181. Available online: https://pubmed.ncbi.nlm.nih.gov/33189883/ (accessed on 22 March 2022). [CrossRef]
- Darbinian, N.; Selzer, M.E. Oligodendrocyte pathology in fetal alcohol spectrum disorders. Neural Regen. Res. 2022, 17, 497–502. Available online: https://pubmed.ncbi.nlm.nih.gov/34380877/ (accessed on 22 March 2022).
- Mooney, S.M.; Miller, M.W. Time-specific effects of ethanol exposure on cranial nerve nuclei: Gastrulation and neuronogenesis. Exp. Neurol. 2007, 205, 56–63. Available online: https://pubmed.ncbi.nlm.nih.gov/17320867/ (accessed on 22 March 2022). [CrossRef] [Green Version]
- Sulik, K.K.; Johnston, M.C.; Webb, M.A. Fetal alcohol syndrome: Embryogenesis in a mouse model. Science 1981, 214, 936–938. Available online: https://science.sciencemag.org/content/214/4523/936 (accessed on 23 June 2022). [CrossRef] [PubMed]
- Sulik, K.K.; Lauder, J.M.; Dehart, D.B. Brain malformations in prenatal mice following acute maternal ethanol administration. Int. J. Dev. Neurosci. 1984, 2, 203–214. [Google Scholar] [CrossRef] [PubMed]
- Sulik, K.K.; Cook, C.S.; Webster, W.S. Teratogens and craniofacial malformations: Relationships to cell death. Development 1988, 103, 213–232. [Google Scholar] [CrossRef] [PubMed]
- Sulik, K.K. Genesis of Alcohol-Induced Craniofacial Dysmorphism. Exp. Biol. Med. 2005, 230, 366–375. [Google Scholar] [CrossRef] [PubMed]
- Sulik, K.K. Critical Periods for Alcohol Teratogenesis in Mice, with Special Reference to the Gastrulation Stage of Embryogenesis. Ciba Found. Symp. 2008, 105, 124–141. [Google Scholar]
- Godin, E.A.; O’Leary-Moore, S.K.; Khan, A.A.; Parnell, S.E.; Ament, J.J.; Dehart, D.B.; Johnson, B.W.; Allan Johnson, G.; Styner, M.A.; Sulik, K.K. Magnetic resonance microscopy defines ethanol-induced brain abnormalities in prenatal mice: Effects of acute insult on gestational day 7. Alcohol. Clin. Exp. Res. 2010, 34, 98–111. Available online: https://www.ncbi.nlm.nih.gov/pubmed/19860813/ (accessed on 6 May 2022). [CrossRef]
- Sulik, K.K.; Johnston, M.C.; Daft, P.A.; Russell, W.E.; Dehart, D.B.; Opitz, J.M.; Reynolds, J.F. Fetal alcohol syndrome and DiGeorge anomaly. Critical ethanol exposure periods for craniofacial malformations as illustrated in an animal model. Am. J. Med. Genet. 1986, 25 (Suppl. S2), 97–112. [Google Scholar] [CrossRef]
- Samson, H.H.; Grant, K.A. Ethanol-induced microcephaly in neonatal rats: Relation to dose. Alcohol. Clin. Exp. Res. 1984, 8, 201–203. Available online: https://pubmed.ncbi.nlm.nih.gov/6375432/ (accessed on 23 March 2022). [CrossRef]
- Tufan, A.C.; Abban, G.; Akdogan, I.; Erdogan, D.; Ozogul, C. The effect of in ovo ethanol exposure on retina and optic nerve in a chick embryo model system. Reprod. Toxicol. 2007, 23, 75–82. [Google Scholar] [CrossRef]
- Kashyap, B.; Frederickson, L.C.; Stenkamp, D.L. Mechanisms for persistent microphthalmia following ethanol exposure during retinal neurogenesis in zebrafish embryos. Vis. Neurosci. 2007, 24, 409–421. [Google Scholar] [CrossRef] [Green Version]
- O’Neil, E. Effects of Prenatal Alcohol Exposure on Ocular Development|The Embryo Project Encyclopedia. embryo.asu.edu. 2011. Available online: https://embryo.asu.edu/pages/effects-prenatal-alcohol-exposure-ocular-development (accessed on 23 March 2022).
- Harris, S.J.; Wilce, P.; Bedi, K.S. Exposure of rats to a high but not low dose of ethanol during early postnatal life increases the rate of loss of optic nerve axons and decreases the rate of myelination. J. Anat. 2000, 197, 477–485. [Google Scholar] [CrossRef] [Green Version]
- Amini, S.; Merabova, N.; Khalili, K.; Darbinian, N. p38SJ, a novel DINGG protein protects neuronal cells from alcohol induced injury and death. J. Cell. Physiol. 2009, 221, 499–504. Available online: https://pubmed.ncbi.nlm.nih.gov/19739100/ (accessed on 22 March 2022). [CrossRef] [PubMed] [Green Version]
- Mercer, K.E.; Hennings, L.; Ronis, M.J. Alcohol consumption, Wnt/β-catenin signaling, and hepatocarcinogenesis. Adv. Exp. Med. Biol. 2015, 815, 185–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goetzl, L.; Darbinian, N.; Goetzl, E.J. Novel window on early human neurodevelopment via fetal exosomes in maternal blood. Ann. Clin. Transl. Neurol. 2016, 3, 381–385. Available online: https://pubmed.ncbi.nlm.nih.gov/27231707/ (accessed on 22 March 2022). [CrossRef] [PubMed]
- Goetzl, L.; Darbinian, N.; Merabova, N. Noninvasive assessment of fetal central nervous system insult: Potential application to prenatal diagnosis. Prenat. Diagn. 2019, 39, 609–615. [Google Scholar] [CrossRef]
- Ramsey, P.H. Critical Values for Spearman’s Rank Order Correlation. J. Educ. Stat. 1989, 14, 245–253. Available online: https://eric.ed.gov/?id=EJ403288 (accessed on 23 March 2022).
- Fowler, J.; Cohen, L.; Jarvis, P. Practical Statistics for Field Biology, 2nd ed.; John Wiley & Sons: Chichester, UK, 2009; p. 132. [Google Scholar]
- Rosett, H.L. A Clinical Perspective of the Fetal Alcohol Syndrome. Alcohol. Clin. Exp. Res. 1980, 4, 119–122. [Google Scholar] [CrossRef] [PubMed]
- Abdelrahman, A.; Conn, R. Eye abnormalities in fetal alcohol syndrome. Ulst. Med. J. 2009, 78, 164–165. Available online: https://pubmed.ncbi.nlm.nih.gov/19907681/ (accessed on 22 March 2022).
- Strömland, K. Visual impairment and ocular abnormalities in children with fetal alcohol syndrome. Addict. Biol. 2004, 9, 153–157. [Google Scholar] [CrossRef]
- Rice, D.; Barone, S. Critical periods of vulnerability for the developing nervous system: Evidence from humans and animal models. Environ. Health Perspect. 2000, 108 (Suppl. 3), 511–533. [Google Scholar]
- Bayer, S.A.; Altman, J. Directions in neurogenetic gradients and patterns of anatomical connections in the telencephalon. Prog. Neurobiol. 1987, 29, 57–106. [Google Scholar] [CrossRef] [PubMed]
- Strömland, K. Ocular abnormalities in the fetal alcohol syndrome. Acta Ophthalmol. 1985, 171, 1–50. [Google Scholar]
- Strömland, K. Ocular involvement in the fetal alcohol syndrome. Surv. Ophthalmol. 1987, 31, 277–284. Available online: https://www.sciencedirect.com/science/article/pii/0039625787900282 (accessed on 7 March 2019). [CrossRef] [PubMed]
- Velázquez-Marrero, C.; Burgos, A.; García, J.O.; Palacio, S.; Marrero, H.G.; Bernardo, A.; Pérez-Laspiur, J.; Rivera-Oliver, M.; Seale, G.; Treistman, S.N. Alcohol Regulates BK Surface Expression via Wnt/β-Catenin Signaling. J. Neurosci. Off. J. Soc. Neurosci. 2016, 36, 10625–10639. Available online: https://pubmed.ncbi.nlm.nih.gov/27733613/ (accessed on 23 March 2022). [CrossRef] [PubMed] [Green Version]
- Flentke, G.R.; Garic, A.; Amberger, E.; Hernandez, M.; Smith, S.M. Calcium-mediated repression of β-catenin and its transcriptional signaling mediates neural crest cell death in an avian model of fetal alcohol syndrome. Birth Defects Res. Part A Clin. Mol. Teratol. 2011, 91, 591–602. [Google Scholar] [CrossRef] [Green Version]
- Pinazo-Durán, M.D. Efecto de la Exposicion pre y Postnatal al Alcohol Sobre el Desarrollo del Nervio Optico, en la Rata. dialnet.unirioja.es. 1991. Available online: https://dialnet.unirioja.es/servlet/tesis?codigo=227382 (accessed on 23 March 2022).
- Parson, S.H.; Dhillon, B.; Findlater, G.S.; Kaufman, M.H. Optic nerve hypoplasia in the fetal alcohol syndrome: A mouse model. J. Anat. 1995, 186 Pt 2, 313–320. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1167188/ (accessed on 23 March 2022).
- Phillips, D.E.; Krueger, S.K. Effects of combined pre- and postnatal ethanol exposure (three trimester equivalency) on glial cell development in rat optic nerve. Int. J. Dev. Neurosci. Off. J. Int. Soc. Dev. Neurosci. 1992, 10, 197–206. Available online: https://pubmed.ncbi.nlm.nih.gov/1442168/ (accessed on 23 March 2022). [CrossRef]
- Samorajski, T.; Lancaster, F.; Wiggins, R.C. Fetal ethanol exposure: A morphometric analysis of myelination in the optic nerve. Int. J. Dev. Neurosci. Off. J. Int. Soc. Dev. Neurosci. 1986, 4, 369–374. Available online: https://pubmed.ncbi.nlm.nih.gov/3455596/ (accessed on 23 March 2022). [CrossRef]
- Chiappelli, F.; Taylor, A.N.; Espinosa de los Monteros, A.; de Vellis, J. Fetal alcohol delays the developmental expression of myelin basic protein and transferrin in rat primary oligodendrocyte cultures. Int. J. Dev. Neurosci. Off. J. Int. Soc. Dev. Neurosci. 1991, 9, 67–75. Available online: https://pubmed.ncbi.nlm.nih.gov/1707580/ (accessed on 22 March 2022). [CrossRef]
- Zoeller, R.T.; Butnariu, O.V.; Fletcher, D.L.; Riley, E.P. Limited postnatal ethanol exposure permanently alters the expression of mRNAS encoding myelin basic protein and myelin-associated glycoprotein in cerebellum. Alcohol. Clin. Exp. Res. 1994, 18, 909–916. Available online: https://pubmed.ncbi.nlm.nih.gov/7526726/ (accessed on 23 March 2022). [CrossRef]
- Bichenkov, E.; Ellingson, J.S. Ethanol alters the expressions of c-Fos and myelin basic protein in differentiating oligodendrocytes. Alcohol 2009, 43, 627–634. Available online: https://pubmed.ncbi.nlm.nih.gov/20004340/ (accessed on 22 March 2022). [CrossRef]
- Snipes, G.J.; Suter, U.; Welcher, A.A.; Shooter, E.M. Characterization of a novel peripheral nervous system myelin protein (PMP-22/SR13). J. Cell Biol. 1992, 117, 225–238. Available online: https://rupress.org/jcb/article/117/1/225/14350 (accessed on 26 February 2022). [CrossRef]
- Parmantier, E.; Braun, C.; Thomas, J.L.; Peyron, F.; Martinez, S.; Zalc, B. PMP-22 expression in the central nervous system of the embryonic mouse defines potential transverse segments and longitudinal columns. J. Comp. Neurol. 1997, 378, 159–172. Available online: https://pubmed.ncbi.nlm.nih.gov/9120057/ (accessed on 23 March 2022). [CrossRef]
- Hagedorn, L.; Suter, U.; Sommer, L. P0 and PMP22 mark a multipotent neural crest-derived cell type that displays community effects in response to TGF-beta family factors. Development 1999, 126, 3781–3794. Available online: https://pubmed.ncbi.nlm.nih.gov/10433908/ (accessed on 22 March 2022). [CrossRef] [PubMed]
- Parmantier, E.; Cabon, F.; Braun, C.; D’Urso, D.; Müller, H.W.; Zalc, B. Peripheral myelin protein-22 is expressed in rat and mouse brain and spinal cord motoneurons. Eur. J. Neurosci. 1995, 7, 1080–1088. Available online: https://pubmed.ncbi.nlm.nih.gov/7613613/ (accessed on 23 March 2022). [CrossRef]
- De Gasperi, R.; Sosa, M.; Naumowicz, Z.; Hof, P.; Notterpek, L.; Davis, K.; Buxbaum, J.D.; Elder, G.A. Peripheral myelin protein-22 is expressed in CNS myelin. Transl. Neurosci. 2010, 1, 282–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chanson, J.B.; Echaniz-Laguna, A.; Blanc, F.; Lacour, A.; Ballonzoli, L.; Kremer, S.; Namer, I.J.; Lannes, B.; Tranchant, C.; Vermersch, P.; et al. Central nervous system abnormalities in patients with PMP22 gene mutations: A prospective study. J. Neurol. Neurosurg. Psychiatry 2013, 84, 392–397. Available online: https://pubmed.ncbi.nlm.nih.gov/23243264/ (accessed on 22 March 2022). [CrossRef] [PubMed]
- Brandt, A.U.; Meinert-Bohn, E.; Rinnenthal, J.L.; Zimmermann, H.; Mikolajczak, J.; Oberwahrenbrock, T.; Papazoglou, S.; Pfüller, C.F.; Schinzel, J.; Tackenberg, B.; et al. Afferent Visual Pathway Affection in Patients with PMP22 Deletion-Related Hereditary Neuropathy with Liability to Pressure Palsies. PLoS ONE 2016, 11, e0164617. Available online: https://pubmed.ncbi.nlm.nih.gov/27749933/ (accessed on 22 March 2022). [CrossRef] [PubMed] [Green Version]
- Conrad, A.H.; Albrecht, M.; Pettit-Scott, M.; Conrad, G.W. Embryonic Corneal Schwann Cells Express Some Schwann Cell Marker mRNAs, but No Mature Schwann Cell Marker Proteins. Investig. Opthalmol. Vis. Sci. 2009, 50, 4173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bennett, J.L.; Zeiler, S.R.; Jones, K.R. Patterned expression of BDNF and NT-3 in the retina and anterior segment of the developing mammalian eye. Investig. Ophthalmol. Vis. Sci. 1999, 40, 2996–3005. Available online: https://pubmed.ncbi.nlm.nih.gov/10549663/ (accessed on 22 March 2022).
- Brennan, D.; Giles, S. Ocular Involvement in Fetal Alcohol Spectrum Disorder: A Review. Curr. Pharm. Des. 2014, 20, 5377–5387. [Google Scholar] [CrossRef] [PubMed]
- Tao, F.; Soffers, J.; Hu, D.; Chen, S.; Gao, X.; Zhang, Y.; Zhao, C.; Smith, S.E.; Unruh, J.R.; Zhang, D.; et al. β-Catenin and Associated Proteins Regulate Lineage Differentiation in Ground State Mouse Embryonic Stem Cells. Stem Cell Rep. 2020, 15, 662–676. Available online: https://pubmed.ncbi.nlm.nih.gov/32822591/ (accessed on 23 March 2022). [CrossRef]
- Kemler, R.; Hierholzer, A.; Kanzler, B.; Kuppig, S.; Hansen, K.; Taketo, M.M.; de Vries, W.N.; Knowles, B.B.; Solter, D. Stabilization of β-catenin in the mouse zygote leads to premature epithelial-mesenchymal transition in the epiblast. Development 2004, 131, 5817–5824. [Google Scholar] [CrossRef]
- Fujimura, N. WNT/β-Catenin Signaling in Vertebrate Eye Development. Front. Cell Dev. Biol. 2016, 30, 4. [Google Scholar] [CrossRef] [Green Version]
- Rudloff, S.; Kemler, R. Differential requirements for b-catenin during mouse development. Development 2012, 139, 3711–3721. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.; Pang, K.M.; Evans, M.; Hay, E.D. Overexpression of beta-catenin induces apoptosis independent of its transactivation function with LEF-1 or the involvement of major G1 cell cycle regulators. Mol. Biol. Cell 2000, 11, 3509–3523. Available online: https://pubmed.ncbi.nlm.nih.gov/11029052/ (accessed on 22 March 2022). [CrossRef]
- Goetzl, L.; Thompson-Felix, T.; Darbinian, N.; Merabova, N.; Merali, S.; Merali, C.; Sanserino, K.; Tatevosian, T.; Fant, B.; Wimmer, M.E. Novel biomarkers to assess in utero effects of maternal opioid use: First steps toward understanding short- and long-term neurodevelopmental sequelae. Genes Brain Behav. 2019, 18, e12583. Available online: https://pubmed.ncbi.nlm.nih.gov/31119847/ (accessed on 22 March 2022). [CrossRef]
- Dukes, K.A.; Burd, L.; Elliott, A.J.; Fifer, W.P.; Folkerth, R.D.; Hankins, G.D.; Hereld, D.; Hoffman, H.J.; Myers, M.M.; Odendaal, H.J.; et al. The Safe Passage Study: Design, Methods, Recruitment, and Follow-Up Approach. Paediatr. Perinat. Epidemiol. 2014, 28, 455–465. [Google Scholar] [CrossRef] [Green Version]
- Stewart, S.H.; Koch, D.G.; Burgess, D.M.; Willner, I.R.; Reuben, A. Sensitivity and specificity of urinary ethyl glucuronide and ethyl sulfate in liver disease patients. Alcohol Clin. Exp. Res. 2013, 37, 150–155. [Google Scholar] [CrossRef]
- Spong, C.Y.; Mercer, B.M.; D’Alton, M.; Kilpatrick, S.; Blackwell, S.; Saade, G. Timing of indicated late-preterm and early-term birth. Obstet. Gynecol. 2011, 118 Pt 1, 323–333. [Google Scholar] [CrossRef] [Green Version]
- Darbinyan, A.; Kaminski, R.; White, M.K.; Darbinian-Sarkissian, N.; Khalili, K. Polyomavirus JC infection inhibits differentiation of oligodendrocyte progenitor cells. J. Neurosci. Res. 2012, 91, 116–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosner, B. Fundamentals of Biostatistics, 7th ed.; Brooks/Cole: Boston, MA, USA, 2011. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Qiagen Data Analysis Handbook. RT2 Profiler PCR Array Handbook. HB-1011-007 06/2019. 2019. Available online: www.qiagen.com (accessed on 1 June 2019).
- Institute of Medicine (US) Committee on Assessing Genetic Risks; Andrews, L.B.; Fullarton, J.E.; Holtzman, N.A.; Motulsky, A.G. (Eds.) Assessing Genetic Risks: Implications for Health and Social Policy; Social, Legal, and Ethical Implications of Genetic Testing; National Academies Press (US): Washington, DC, USA, 1994; p. 8. Available online: https://www.ncbi.nlm.nih.gov/books/NBK236044/ (accessed on 1 April 1994).








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gestation Period in Weeks | 1 | 4 | 7 | 10 | 13 | 16 | 19 | 22 | 28 | 31 | 34 | 37 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Alcohol | |||||||||||||
| Brain |  | ||||||||||||
| Heart |  | ||||||||||||
| Arms |  | ||||||||||||
| Eyes |  | ||||||||||||
| Legs |  | ||||||||||||
| Teeth |  | ||||||||||||
| Palate |  | ||||||||||||
| Ears |  | ||||||||||||
| Genital Area |  | ||||||||||||
| Controls (n = 20) | EtOH Users (n = 20) | |
|---|---|---|
| Maternal age (years ±SD) | 24.15 ± 2.3 | 28.0 ± 2.7 |
| Parity (±SD) | 0.83 ± 0.42 | 0.82 ± 0.67 |
| GA (weeks ±SD) | 14.92 ± 1.58 | 15.22 ± 1.6 |
| GA range (weeks) | 11.1 to 21 | 11.5 to 21 |
| Body Mass Index (±SD) | 24.4 ± 3.3 | 26.1 ± 1.5 |
| Hispanic Ethnicity (%) | 15 | 15 |
| Race | ||
| White (%) | 50 | 50 |
| Black (%) | 50 | 50 |
| Tobacco Use (%) | 20 | 25 |
| Male Fetal Sex (%) | 50 | 50 |
| Control, GA 11-14 (n = 10, ±SD) | Control, GA 14.1-21 (n = 10, ±SD) | EtOH, GA 11-14 (n = 10, ±SD) | EtOH, GA 14.1-21 (n = 10, ±SD) | |
|---|---|---|---|---|
| Eye Diameter (wet specimen in PFA), mm | 5.8 ± 0.8; p < 0.05 | 10.2 ± 1.5; p = 0.055 (n.s.) | 5.0 ± 0.6; p < 0.05 | 9.5 ± 1.7; p = 0.060 (n.s.) |
| Eye Diameter (histological sections in OCT), mm | 5.4 ± 0.9; p < 0.05 | 9.9 ± 0.8; p < 0.05 | 4.9 ± 0.7; p < 0.05 | 9.0 ± 0.9; p < 0.05 |
| Pupil Diameter (wet specimen in PFA), mm | 5.3 ± 0.4; p < 0.05 | 4.6 ± 0.7; p < 0.05 |
| Race | White | African American | |||
|---|---|---|---|---|---|
| 1st Trimester (mm) | 2nd Trimester (mm) | 1st Trimester (mm) | 2nd Trimester (mm) | ||
| EtOH Exposure | |||||
| Control (n = 20) | 4.1 ± 0.2 | 7.4 ± 0.5 | 3.52 ± 0.1 | 6.72 ± 0.4 | |
| EtOH (n = 20) | 3.34 ± 0.3; p = 0.007 | 6.76 ± 0.6; p = 0.05 | 2.26 ± 0.2; p = 0.0005 | 7.42 ± 0.6; p = 0.05 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Darbinian, N.; Darbinyan, A.; Sinard, J.; Tatevosian, G.; Merabova, N.; D’Amico, F.; Khader, T.; Bajwa, A.; Martirosyan, D.; Gawlinski, A.K.; et al. Molecular Markers in Maternal Blood Exosomes Allow Early Detection of Fetal Alcohol Spectrum Disorders. Int. J. Mol. Sci. 2023, 24, 135. https://doi.org/10.3390/ijms24010135
Darbinian N, Darbinyan A, Sinard J, Tatevosian G, Merabova N, D’Amico F, Khader T, Bajwa A, Martirosyan D, Gawlinski AK, et al. Molecular Markers in Maternal Blood Exosomes Allow Early Detection of Fetal Alcohol Spectrum Disorders. International Journal of Molecular Sciences. 2023; 24(1):135. https://doi.org/10.3390/ijms24010135
Chicago/Turabian StyleDarbinian, Nune, Armine Darbinyan, John Sinard, Gabriel Tatevosian, Nana Merabova, Faith D’Amico, Tarek Khader, Ahsun Bajwa, Diana Martirosyan, Alina K. Gawlinski, and et al. 2023. "Molecular Markers in Maternal Blood Exosomes Allow Early Detection of Fetal Alcohol Spectrum Disorders" International Journal of Molecular Sciences 24, no. 1: 135. https://doi.org/10.3390/ijms24010135